Homo (du latin homō, signifiant » homme « ) est le genre apparu dans le genre Australopithecus, par ailleurs éteint, qui englobe l’espèce existante Homo sapiens (les humains modernes), plus plusieurs espèces éteintes classées comme ancestrales ou étroitement liées aux humains modernes (selon l’espèce), notamment Homo erectus et Homo neanderthalensis.Le genre est considéré comme apparu avec l’apparition de l’Homo habilis, il y a un peu plus de deux millions d’années. Le genre Homo, avec le genre Paranthropus est probablement frère de A. africanus dans le genre Australopithecus, qui lui-même s’était auparavant séparé de la lignée de Pan, les chimpanzés.
Homo erectus est apparu il y a environ deux millions d’années et, au cours de plusieurs migrations précoces, il s’est répandu en Afrique (où il est surnommé Homo ergaster) et en Eurasie. C’était probablement la première espèce humaine à vivre dans une société de chasseurs-cueilleurs et à maîtriser le feu. Espèce adaptative et performante, Homo erectus a persisté pendant plus d’un million d’années et a progressivement divergé vers de nouvelles espèces il y a environ 500 000 ans.
Homo sapiens (les humains anatomiquement modernes) est apparu il y a près de 300 000 à 200 000 ans, très probablement en Afrique, et Homo neanderthalensis est apparu à peu près au même moment en Europe et en Asie occidentale.L’H. sapiens s’est dispersé d’Afrique en plusieurs vagues, peut-être dès 250 000 ans et certainement avant 130 000 ans, la dispersion dite australe commençant il y a environ 70-50 000 ans et conduisant à la colonisation durable de l’Eurasie et de l’Océanie il y a 50 000 ans. Des espèces humaines archaïques (non sapiens) distinctes auraient survécu jusqu’à il y a environ 40 000 ans (extinction de Neandertal), avec une possible survie tardive d’espèces hybrides jusqu’à il y a 12 000 ans (peuple de la grotte de Red Deer).
Noms et taxonomie

Tableau de l’arbre évolutif mettant en avant la sous-famille Homininae et la tribu Hominini. Après avoir divergé de la lignée des Ponginae, les premiers Homininae se sont divisés en tribus Hominini et Gorillini. Les premiers Hominini se sont encore divisés, séparant la lignée d’Homo de la lignée de Pan. Actuellement, la tribu Hominini désigne les sous-tribus Hominina, contenant le genre Homo ; Panina, le genre Pan ; et Australopithecina, avec plusieurs genres éteints – les sous-tribus ne sont pas étiquetées sur ce graphique.
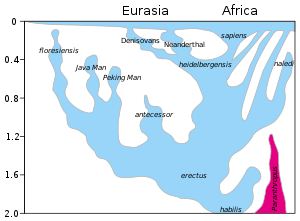
Un modèle de l’évolution du genre Homo au cours des 2 derniers millions d’années (axe vertical). L’expansion rapide « Out of Africa » de H. sapiens est indiquée en haut du diagramme, avec le mélange indiqué avec les Néandertaliens, les Denisovans et des hominines africains archaïques non spécifiés. La survie tardive des australopithèques robustes (Paranthropus) aux côtés d’Homo jusqu’à 1,2 Mya est indiquée en violet.
Articles principaux : Taxonomie humaine, Noms de l’espèce humaine Voir Homininae pour un aperçu de la taxonomie.
Le nom latin homō (génitif hominis) signifie « être humain » ou « homme » au sens générique de « être humain, humanité ». Le nom binomial Homo sapiens a été inventé par Carl Linnaeus (1758). Les noms des autres espèces du genre ont été introduits à partir de la seconde moitié du XIXe siècle (H. neanderthalensis 1864, H. erectus 1892).
Même aujourd’hui, le genre Homo n’a pas été strictement défini. Depuis que les premiers fossiles humains ont commencé à émerger lentement de la terre, les frontières et les définitions du genre Homo ont été mal définies et en constante évolution. Parce qu’il n’y avait aucune raison de penser qu’il aurait un jour d’autres membres, Carl Linnaeus n’a même pas pris la peine de définir Homo lorsqu’il l’a créé pour la première fois pour les humains au 18e siècle. La découverte de Neandertal a apporté le premier ajout.
Le genre Homo a reçu son nom taxonomique pour suggérer que les espèces qui le composent peuvent être classées comme humaines. Et, au cours des décennies du 20ème siècle, les découvertes de fossiles d’espèces pré-humaines et humaines précoces de la fin du Miocène et du début du Pliocène ont produit un riche mélange pour débattre des classifications. Le débat se poursuit sur la distinction entre Homo et Australopithecus – ou, en fait, entre Homo et Pan, puisqu’un groupe de scientifiques soutient que les deux espèces de chimpanzés devraient être classées dans le genre Homo plutôt que Pan. Malgré cela, classer les fossiles d’Homo coïncide avec les preuves de : (1) la bipédie humaine compétente chez Homo habilis héritée de l’Australopithèque antérieur d’il y a plus de quatre millions d’années, comme le démontrent les empreintes de Laetoli ; et (2) la culture des outils humains ayant commencé il y a 2,5 millions d’années.
De la fin du XIXe siècle au milieu du XXe siècle, un certain nombre de nouveaux noms taxonomiques, y compris de nouveaux noms génériques, ont été proposés pour les premiers fossiles humains ; la plupart ont depuis été fusionnés avec Homo, en reconnaissance du fait que Homo erectus était une espèce unique et singulière avec une grande étendue géographique des premières migrations. Beaucoup de ces noms sont maintenant surnommés « synonymes » avec Homo, notamment Pithecanthropus,Protanthropus,Sinanthropus,Cyphanthropus,Africanthropus,Telanthropus,Atlanthropus,et Tchadanthropus.
La classification du genre Homo en espèces et sous-espèces est sujette à des informations incomplètes et reste mal faite. Cela a conduit à utiliser des noms communs (« Neanderthal » et « Denisovan »), même dans les articles scientifiques, pour éviter les noms trinomiaux ou l’ambiguïté de classer les groupes comme incertae sedis (placement incertain) – par exemple, H. neanderthalensis vs H. sapiens neanderthalensis, ou H. georgicus vs H. erectus georgicus. Certaines espèces récemment éteintes du genre Homo n’ont été découvertes que récemment et n’ont pas encore de nom binomial consensuel (voir l’hominine de Denisova et l’homme de la grotte de Red Deer). Depuis le début de l’Holocène, il est probable qu’Homo sapiens (humains anatomiquement modernes) soit la seule espèce d’Homo existante.
John Edward Gray (1825) a été l’un des premiers partisans de la classification des taxons en désignant des tribus et des familles.Wood et Richmond (2000) ont proposé que Hominini ( » hominines « ) soit désigné comme une tribu comprenant toutes les espèces d’humains primitifs et de pré-humains ancestraux aux humains jusqu’après le dernier ancêtre commun chimpanzé-homme ; et qu’Hominina soit désignée comme une sous-tribu d’Hominini pour n’inclure que le genre Homo, c’est-à-dire sans inclure les premiers hominines marchant debout du Pliocène tels que Australopithecus, Orrorin tugenensis, Ardipithecus, ou Sahelanthropus.Des désignations alternatives à Hominina ont existé, ou ont été proposées : Australopithecinae (Gregory & Hellman 1939) et Preanthropinae (Cela-Conde & Altaba 2002) ;et plus tard, Cela-Conde et Ayala (2003) ont proposé que les quatre genres Australopithecus, Ardipithecus, Praeanthropus et Sahelanthropus soient regroupés avec Homo au sein de Hominini (sans pan).
Évolution
Informations complémentaires : Chronologie de l’évolution humaine Voir Hominini et Chimpanzé-dernier ancêtre commun de l’homme pour la séparation entre Australopithecina et Panina.
Australopithecus
Plus d’informations : Australopithèque
Reconstruction médico-légale de A. afarensis
Plusieurs espèces, dont Australopithecus garhi, Australopithecus sediba, Australopithecus africanus et Australopithecus afarensis, ont été proposées comme ancêtre direct ou sœur de la lignée Homo.Ces espèces ont des caractéristiques morphologiques qui les alignent avec Homo, mais il n’y a pas de consensus pour savoir laquelle a donné naissance à Homo.
En particulier depuis les années 2010, la délimitation d’Homo dans les Australopithèques est devenue plus controversée. Traditionnellement, on considère que l’avènement d’Homo coïncide avec la première utilisation d’outils en pierre (l’industrie Oldowan), et donc par définition avec le début du Paléolithique inférieur.Mais en 2010, des preuves ont été présentées qui semblent attribuer l’utilisation d’outils en pierre à Australopithecus afarensis il y a environ 3,3 millions d’années, soit près d’un million d’années avant la première apparition d’Homo.LD 350-1, un fragment de mandibule fossile daté de 2.8 Mya, découvert en 2015 à Afar, en Éthiopie, a été décrit comme combinant « des traits primitifs observés chez les premiers australopithèques avec une morphologie dérivée observée chez les Homo ultérieurs.Certains auteurs pousseraient le développement d’Homo près ou même au-delà de 3 Mya.D’autres ont émis des doutes quant à l’inclusion d’Homo habilis dans Homo, proposant plutôt une origine d’Homo avec Homo erectus à environ 1,9 Mya.
Le développement physiologique le plus saillant entre les espèces australopithèques antérieures et Homo est l’augmentation du volume endocrânien (VEC), qui passe d’environ 460 cm3 (28 cu in) chez A. garhi à 660 cm3 (40 cu in) chez H. habilis, puis à 760 cm3 (46 cu in) chez H. erectus, 1 250 cm3 (76 cu in) chez H. heidelbergensis et jusqu’à 1 760 cm3 (107 cu in) chez H. neanderthalensis.Cependant, une augmentation régulière de la capacité crânienne est observée déjà chez Autralopithecina et ne s’arrête pas après l’émergence d’Homo, de sorte qu’elle ne sert pas de critère objectif pour définir l’émergence du genre.
Homo habilis
Reconstruction médico-légale d’Homo habilis, exposition au LWL-Museum für Archäologie, Herne, Allemagne (photographie de 2007).
Homo habilis est apparu vers 2,1 Mya. Déjà avant 2010, il a été suggéré que H. habilis ne devrait pas être placé en dehors du genre Homo dans le plus large Australopithecus.La principale raison d’inclure H. habilis dans Homo, son utilisation incontestée d’outils, est devenue obsolète avec la découverte de l’utilisation d’outils d’Australopithecus au moins un million d’années avant H. En outre, H. habilis a longtemps été considéré comme l’ancêtre de l’Homo ergaster (Homo erectus), plus gracile. En 2007, on a découvert que H. habilis et H. erectus ont coexisté pendant une période considérable, ce qui suggère que H. erectus ne dérive pas immédiatement de l’Homo ergaster. Avec la publication du crâne 5 de Dmanisi en 2013, il est devenu moins certain que l’H. erectus asiatique soit un descendant de l’H. ergaster africain, qui était à son tour dérivé de l’H. habilis.Au lieu de cela, H. ergaster et H. erectus semblent être des variantes de la même espèce, qui peut avoir été originaire soit de l’Afrique, soit de l’Asie et largement dispersée dans toute l’Eurasie (y compris l’Europe, l’Indonésie, la Chine) vers 0,5 Mya.
Homo erectus
Articles principaux : Homo erectus
Homo erectus a souvent été supposé s’être développé anagénétiquement à partir d’Homo habilis à partir d’environ 2 millions d’années.Ce scénario a été renforcé avec la découverte d’Homo erectus georgicus,les premiers spécimens d’H. erectus trouvés dans le Caucase, qui semblaient présenter des traits transitionnels avec H. habilis. Sur la base des fossiles de la formation de Koobi Fora, à l’est du lac Turkana au Kenya, Spoor et al. (2007) ont avancé que H. habilis avait pu survivre après l’émergence de H. erectus, de sorte que l’évolution de H. erectus n’aurait pas été anagénétique et que H. erectus aurait existé aux côtés de H. habilis pendant environ un demi-million d’années. habilis pendant environ un demi-million d’années (il y a 1,9 à 1,4 million d’années), au début du Calabre.
Une espèce sud-africaine distincte, Homo gautengensis, a été postulée comme contemporaine d’Homo erectus en 2010.
Phylogénie
Une taxonomie de l’Homo au sein des grands singes est évaluée comme suit, avec Paranthropus et Homo émergeant au sein d’Australopithecus (représenté ici de manière cladistique accordant Paranthropus, Kenyanthropus, et Homo). La phylogénie exacte au sein des australopithèques est encore très controversée. Les dates approximatives de radiation des clades filles sont indiquées en millions d’années (Mya). Graecopithecus, Sahelanthropus, Orrorin, qui sont peut-être des sœurs des australopithèques, ne sont pas représentés ici. Notez que la dénomination des groupements est parfois confuse car souvent certains groupements sont présumés avant la réalisation d’analyses cladistiques.
| Hominoidea |
|
|||||||||||||||||||||||||
| (20.4 Mya) |
| Australopithèques |
Plusieurs des lignées Homo semblent avoir une descendance survivante par introgression dans d’autres lignées. Une lignée archaïque se séparant des autres lignées humaines il y a 1,5 million d’années, peut-être H. erectus, pourrait s’être croisée avec les Denisovans il y a environ 55 000 ans. L’Homo erectus s.s. a survécu jusqu’à il y a 27 000 ans, et l’Homo florensiensis, encore plus basique, a survécu jusqu’à il y a 50 000 ans. En outre, un os de cuisse, daté de 14 000 ans, découvert dans une grotte de Maludong (peuple de la grotte de Red Deer) ressemble fortement à des espèces très anciennes comme l’Homo erectus précoce ou la lignée encore plus archaïque, l’Homo habilis, qui vivait il y a environ 1,5 million d’années. Une partie de la lignée semblable à l’Homo erectus, qui a vécu il y a 1,5 million d’années, semble avoir été introduite chez l’homme moderne par les Denisovans, et plus particulièrement chez les Papous et les aborigènes australiens. Il existe des preuves de l’introgression de H. Heidelbergensis dans H. sapiens. Les génomes des humains d’Afrique non subsaharienne montrent ce qui semble être de nombreux événements d’introgression indépendants impliquant les Néandertaliens et, dans certains cas, les Denisovans, il y a environ 45 000 ans. De même, la structure génétique des Africains subsahariens semble indiquer une introgression à partir d’une lignée humaine archaïque distincte, encore non identifiée, telle que H. heidelbergensis. Australopithecus sediba est sur le point d’être renommé Homo sediba en raison de sa position par rapport, par exemple, à Homo habilis et Homo floresiensis. DispersionVoir aussi : Expansions humaines précoces hors d’Afrique, Mélange de l’homme archaïque avec l’homme moderne, Migrations humaines précoces Vers 1,8 million d’années, Homo erectus est présent à la fois en Afrique de l’Est (Homo ergaster) et en Asie occidentale (Homo georgicus).Les ancêtres de l’Homo floresiensis indonésien pourraient avoir quitté l’Afrique encore plus tôt.  Dispersions successives d’Homo erectus (jaune), d’Homo neanderthalensis (ocre) et d’Homo sapiens (rouge). L’Homo erectus et les espèces humaines archaïques apparentées ou dérivées, au cours des 1,5 million d’années suivantes, se répandent en Afrique et en Eurasie (voir : Origine africaine récente des humains modernes). L’Europe est atteinte vers 0,5 Mya par Homo heidelbergensis. Homo neanderthalensis et Homo sapiens se développent après environ 300 kya. Homo naledi est présent en Afrique australe vers 300 kya. H. sapiens peu après sa première émergence s’est répandu dans toute l’Afrique, et en Asie occidentale en plusieurs vagues, peut-être dès 250 kya, et certainement vers 130 kya. En juillet 2019, des anthropologues ont signalé la découverte de restes vieux de 210 000 ans d’un H. sapiens et de 170 000 ans d’un H. neanderthalensis dans la grotte d’Apidima, dans le Péloponnèse, en Grèce, soit plus de 150 000 ans de plus que les précédentes découvertes de H. sapiens en Europe. La plus notable est la Dispersion Sud de H. sapiens vers 60 kya, qui a conduit au peuplement durable de l’Océanie et de l’Eurasie par des humains anatomiquement modernes.H. sapiens s’est croisé avec des humains archaïques à la fois en Afrique et en Eurasie, en Eurasie notamment avec les Néandertaliens et les Denisovans. Parmi les populations existantes d’Homo sapiens, la division temporelle la plus profonde se trouve chez les San d’Afrique australe, estimée à près de 130 000 ans, voire à plus de 300 000 ans.La division temporelle chez les non-africains est de l’ordre de 60 000 ans dans le cas des Australo-Mélanésiens.La division des Européens et des Asiatiques de l’Est est de l’ordre de 50 000 ans, avec des événements de mélange répétés et significatifs dans toute l’Eurasie au cours de l’Holocène. Les espèces humaines archaïques ont peut-être survécu jusqu’au début de l’Holocène (peuple de la grotte de Red Deer), bien qu’elles aient été pour la plupart éteintes ou absorbées par les populations H. sapiens en expansion vers 40 kya (extinction de Neandertal). Liste des lignéesVoir aussi : Liste des fossiles de l’évolution humaine Le statut d’espèce de H. rudolfensis, H. ergaster, H. georgicus, H. antecessor, H. cepranensis, H. rhodesiensis, H. neanderthalensis, hominine de Denisova, peuple de la grotte de Red Deer, et H. floresiensis reste en débat. H. heidelbergensis et H. neanderthalensis sont étroitement liés l’un à l’autre et ont été considérés comme des sous-espèces de H. sapiens. Il y a eu historiquement une tendance à postuler de « nouvelles espèces humaines » basées sur aussi peu qu’un fossile individuel. Une approche « minimaliste » de la taxonomie humaine reconnaît au maximum trois espèces, Homo habilis (2,1-1,5 Mya, appartenance à Homo douteuse), Homo erectus (1,8-0,1 Mya, incluant la majorité de l’âge du genre, et la majorité des variétés archaïques en tant que sous-espèces, y compris H. heidelbergensis en tant que variété tardive ou transitoire) et Homo sapiens (300 kya à aujourd’hui, y compris H. neanderthalensis et d’autres variétés en tant que sous-espèces). Dans ce contexte, le terme « espèce » ne signifie pas nécessairement que l’hybridation et l’introgression étaient impossibles à l’époque. Cependant, il est souvent utilisé comme un terme commode, mais il doit être considéré comme signifiant au mieux une lignée générique, et au pire des groupes. En général, les définitions et la méthodologie des critères de délimitation des « espèces » ne font pas l’objet d’un consensus en anthropologie ou en paléontologie. En effet, les mammifères et peuvent typiquement se croiser pendant 2 à 3 millions d’années ou plus, donc toutes les « espèces » contemporaines d’Homo dans le genre Homo auraient potentiellement pu se croiser à l’époque, et l’introgression depuis l’extérieur du genre Homo ne peut être exclue a priori. Il a été suggéré que H. naledi pourrait avoir été un hybride avec un australipith (pris pour signifier au-delà d’Homo, ndlr) ayant survécu tardivement, malgré le fait que ces lignées sont généralement considérées comme éteintes depuis longtemps. Comme discuté ci-dessus, de nombreuses introgressions ont eu lieu entre les lignées, avec des preuves d’introgression après une séparation de 1,5 Millions d’années. Notes de bas de page
. |
