Les dommages à l’ADNModifier

Les dommages à l’ADN sont considérés comme la principale cause sous-jacente des néoplasmes malins appelés cancers. Son rôle central dans la progression vers le cancer est illustré dans la figure de cette section, dans l’encadré près du sommet (les caractéristiques centrales des dommages à l’ADN, des altérations épigénétiques et de la réparation déficiente de l’ADN dans la progression vers le cancer sont indiquées en rouge). Les lésions de l’ADN sont très courantes. Les lésions de l’ADN d’origine naturelle (principalement dues au métabolisme cellulaire et aux propriétés de l’ADN dans l’eau à la température du corps) se produisent à un rythme de plus de 60 000 nouvelles lésions, en moyenne, par cellule humaine, par jour. Des dommages supplémentaires à l’ADN peuvent résulter de l’exposition à des agents exogènes. La fumée de tabac provoque une augmentation des dommages exogènes à l’ADN, et ces dommages à l’ADN sont la cause probable du cancer du poumon dû au tabagisme. La lumière UV du rayonnement solaire provoque des dommages à l’ADN qui sont importants dans le mélanome. L’infection par Helicobacter pylori produit des niveaux élevés d’espèces réactives de l’oxygène qui endommagent l’ADN et contribuent au cancer gastrique. Les acides biliaires, à des niveaux élevés dans le côlon des humains ayant une alimentation riche en graisses, causent également des dommages à l’ADN et contribuent au cancer du côlon. Katsurano et al. ont indiqué que les macrophages et les neutrophiles dans un épithélium colique enflammé sont la source d’espèces réactives de l’oxygène causant des dommages à l’ADN qui initient la tumorigenèse colique. Certaines sources de dommages à l’ADN sont indiquées dans les encadrés en haut de la figure de cette section.
Les individus présentant une mutation de la lignée germinale entraînant une déficience dans l’un des 34 gènes de réparation de l’ADN (voir article Trouble de la réparation de l’ADN) présentent un risque accru de cancer. Certaines mutations de la lignée germinale dans les gènes de réparation de l’ADN entraînent jusqu’à 100 % de risque de cancer au cours de la vie (par exemple, les mutations p53). Ces mutations de la lignée germinale sont indiquées dans un encadré à gauche de la figure avec une flèche indiquant leur contribution au déficit de réparation de l’ADN.
Environ 70 % des néoplasmes malins n’ont pas de composante héréditaire et sont appelés « cancers sporadiques ». Seule une minorité de cancers sporadiques présente un déficit de réparation de l’ADN dû à une mutation dans un gène de réparation de l’ADN. Cependant, la majorité des cancers sporadiques présentent une déficience dans la réparation de l’ADN due à des altérations épigénétiques qui réduisent ou réduisent au silence l’expression du gène de réparation de l’ADN. Par exemple, sur 113 cancers colorectaux séquentiels, seuls quatre présentaient une mutation faux-sens dans le gène de réparation de l’ADN MGMT, tandis que la majorité présentait une expression réduite de MGMT due à la méthylation de la région promotrice de MGMT (une altération épigénétique). Cinq rapports présentent des preuves qu’entre 40 et 90 % des cancers colorectaux ont une expression réduite de MGMT due à la méthylation de la région promotrice de MGMT.
De même, sur 119 cas de cancers colorectaux déficients en réparation des mésappariements qui manquaient d’expression du gène de réparation de l’ADN PMS2, PMS2 était déficient dans 6 cas en raison de mutations du gène PMS2, tandis que dans 103 cas, l’expression de PMS2 était déficiente parce que son partenaire d’appariement MLH1 était réprimé en raison de la méthylation du promoteur (la protéine PMS2 est instable en l’absence de MLH1). Dans les 10 autres cas, la perte d’expression de PMS2 était probablement due à une surexpression épigénétique du microARN, miR-155, qui régule à la baisse MLH1.
Dans d’autres exemples, des défauts épigénétiques ont été trouvés à des fréquences comprises entre 13 % et 100 % pour les gènes de réparation de l’ADN BRCA1, WRN, FANCB, FANCF, MGMT, MLH1, MSH2, MSH4, ERCC1, XPF, NEIL1 et ATM. Ces défauts épigénétiques sont apparus dans divers cancers (par exemple, sein, ovaire, colorectal et tête et cou). Deux ou trois déficiences de l’expression d’ERCC1, XPF ou PMS2 se produisent simultanément dans la majorité des 49 cancers du côlon évalués par Facista et al. Les altérations épigénétiques provoquant une expression réduite des gènes de réparation de l’ADN est représentée dans un encadré central au troisième niveau à partir du haut de la figure de cette section, et la déficience de réparation de l’ADN qui en résulte est représentée au quatrième niveau.
Lorsque l’expression des gènes de réparation de l’ADN est réduite, les dommages à l’ADN s’accumulent dans les cellules à un niveau plus élevé que la normale, et ces dommages en excès provoquent une augmentation des fréquences de mutation ou d’épimutation. Les taux de mutation augmentent fortement dans les cellules défectueuses dans la réparation des mésappariements de l’ADN ou dans la réparation homologue recombinatoire (HRR).
Lors de la réparation des cassures double brin de l’ADN, ou de la réparation d’autres dommages de l’ADN, les sites de réparation incomplètement dégagés peuvent provoquer un silençage épigénétique des gènes. Les déficiences de la réparation de l’ADN (niveau 4 dans la figure) entraînent une augmentation des dommages à l’ADN (niveau 5 dans la figure) qui se traduit par une augmentation des mutations somatiques et des altérations épigénétiques (niveau 6 dans la figure).
Les défauts de champ, des tissus d’apparence normale présentant de multiples altérations (et discutés dans la section ci-dessous), sont des précurseurs courants du développement du clone de tissu désordonné et proliférant de manière inappropriée dans un néoplasme malin. De tels défauts de champ (deuxième niveau à partir du bas de la figure) peuvent présenter de multiples mutations et altérations épigénétiques.
Une fois qu’un cancer est formé, il présente généralement une instabilité du génome. Cette instabilité est probablement due à une réduction de la réparation de l’ADN ou à des dommages excessifs à l’ADN. En raison de cette instabilité, le cancer continue à évoluer et à produire des sous-clones. Par exemple, un cancer du rein, échantillonné dans 9 zones, présentait 40 mutations ubiquitaires, démontrant l’hétérogénéité de la tumeur (c’est-à-dire présentes dans toutes les zones du cancer), 59 mutations partagées par certaines (mais pas toutes les zones), et 29 mutations « privées » présentes uniquement dans une des zones du cancer.
Défauts de champsModifier
Divers autres termes ont été utilisés pour décrire ce phénomène, notamment « effet de champ », « cancérisation de champ » et « carcinogenèse de champ ». Le terme « cancérisation de champ » a été utilisé pour la première fois en 1953 pour décrire une zone ou un « champ » d’épithélium qui a été préconditionné par des processus (à l’époque) largement inconnus de manière à le prédisposer au développement d’un cancer. Depuis lors, les termes « cancérisation de champ » et « défaut de champ » ont été utilisés pour décrire un tissu pré-malin dans lequel de nouveaux cancers sont susceptibles de se produire.
Les défauts de champ sont importants dans la progression vers le cancer. Cependant, dans la plupart des recherches sur le cancer, comme le souligne Rubin « La grande majorité des études dans la recherche sur le cancer a été réalisée sur des tumeurs bien définies in vivo, ou sur des foyers néoplasiques discrets in vitro. Pourtant, il est prouvé que plus de 80 % des mutations somatiques trouvées dans les tumeurs colorectales humaines à phénotype mutateur se produisent avant le début de l’expansion clonale terminale. De même, Vogelstein et al. soulignent que plus de la moitié des mutations somatiques identifiées dans les tumeurs sont survenues dans une phase prénéoplasique (dans un défaut de champ), pendant la croissance de cellules apparemment normales. De même, les altérations épigénétiques présentes dans les tumeurs peuvent être survenues dans des défauts de champ prénéoplasiques.
Une vision élargie de l’effet de champ a été appelée « effet de champ étiologique », qui englobe non seulement les changements moléculaires et pathologiques dans les cellules prénéoplasiques, mais aussi les influences des facteurs environnementaux exogènes et les changements moléculaires dans le microenvironnement local sur l’évolution néoplasique, de l’initiation de la tumeur au décès du patient.
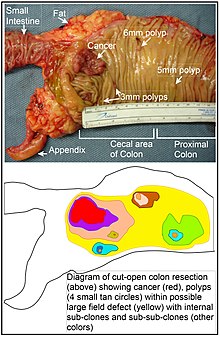
Dans le côlon, un défaut de champ survient probablement par sélection naturelle d’une cellule mutante ou épigénétiquement altérée parmi les cellules souches à la base de l’une des cryptes intestinales sur la surface interne du côlon. Une cellule souche mutante ou épigénétiquement altérée peut remplacer les autres cellules souches voisines par sélection naturelle. Ainsi, une parcelle de tissu anormal peut apparaître. La figure de cette section comprend une photo d’un segment du côlon fraîchement réséqué et ouvert en longueur, montrant un cancer du côlon et quatre polypes. Sous la photo, un diagramme schématique montre comment une grande plaque de cellules mutantes ou épigénétiquement modifiées a pu se former, comme le montre la grande zone en jaune sur le diagramme. À l’intérieur de cette première grande plaque du diagramme (un grand clone de cellules), une deuxième mutation ou altération épigénétique de ce type peut se produire, de sorte qu’une cellule souche donnée acquiert un avantage par rapport aux autres cellules souches de la plaque, et cette cellule souche altérée peut se développer de manière clonale en formant une plaque secondaire, ou sous-clone, à l’intérieur de la plaque initiale. Ceci est indiqué dans le diagramme par quatre petites taches de couleurs différentes dans la grande zone jaune d’origine. Au sein de ces nouvelles taches (sous-clones), le processus peut être répété plusieurs fois, indiqué par les taches encore plus petites au sein des quatre taches secondaires (avec des couleurs encore différentes dans le diagramme) qui s’étendent de manière clonale, jusqu’à ce que des cellules souches apparaissent et forment soit de petits polypes, soit un néoplasme malin (cancer).
Sur la photo, un défaut de champ apparent dans ce segment d’un côlon a généré quatre polypes (étiquetés avec la taille des polypes, 6mm, 5mm, et deux de 3mm, et un cancer d’environ 3 cm de diamètre dans sa dimension la plus longue). Ces néoplasmes sont également indiqués, dans le diagramme sous la photo, par 4 petits cercles de couleur fauve (polypes) et une zone rouge plus grande (cancer). Le cancer sur la photo est apparu dans la région cæcale du côlon, là où le côlon rejoint l’intestin grêle (marqué) et où se trouve l’appendice (marqué). La graisse sur la photo est externe à la paroi extérieure du côlon. Dans le segment de côlon présenté ici, le côlon a été ouvert dans le sens de la longueur afin d’exposer la surface interne du côlon et de montrer le cancer et les polypes qui se produisent dans la paroi épithéliale interne du côlon.
Si le processus général par lequel les cancers sporadiques du côlon apparaissent est la formation d’un clone prénéoplasique qui se propage par sélection naturelle, suivie de la formation de sous-clones internes au sein du clone initial, et de sous-sous-clones à l’intérieur de ceux-ci, alors les cancers du côlon devraient généralement être associés à, et être précédés par, des champs d’anomalie croissante reflétant la succession d’événements prémalins. La région d’anomalie la plus étendue (la zone irrégulière jaune la plus extérieure dans le diagramme) refléterait l’événement le plus précoce dans la formation d’un néoplasme malin.
Dans l’évaluation expérimentale des déficiences spécifiques de réparation de l’ADN dans les cancers, de nombreuses déficiences spécifiques de réparation de l’ADN ont également été montrées pour se produire dans les défauts de champ entourant ces cancers. Le tableau, ci-dessous, donne des exemples pour lesquels il a été démontré que la déficience de réparation de l’ADN dans un cancer était causée par une altération épigénétique, et les fréquences un peu plus faibles avec lesquelles la même déficience de réparation de l’ADN causée par l’épigénétique a été trouvée dans le défaut de champ environnant.
| Cancer | Gène | Fréquence dans le cancer | Fréquence dans le Field Defect | Ref. | Colorectal | MGMT | 46% |
|---|---|---|---|---|
| MGMT | 47% | 47% | 11% | |
| Colorectal | MGMT | 70% | 60% | |
| Colorectal | MSH2 | 13% | 5% | |
| Colorectal | ERCC1 | 100% | 40% | |
| Colorectal | PMS2 | 88% | 50% | |
| Colorectal | XPF | 55% | 40% | Tête et Cou | MGMT | 54% | 38% | Tête et cou | MLH1 | 33% | 25% |
| Tête et cou | MLH1 | 31% | 20% | |
| Estomac | MGMT | 88% | 78% | |
| Estomac | MLH1 | 73% | 20% | |
| Esophage | MLH1 | 77%-100% | 23%-79% |
Certains des petits polypes dans le défaut de champ montré sur la photo du segment de côlon ouvert peuvent être des néoplasmes relativement bénins. Parmi les polypes de moins de 10 mm, découverts lors d’une coloscopie et suivis par des coloscopies répétées pendant 3 ans, 25 % n’ont pas changé de taille, 35 % ont régressé ou diminué de taille tandis que 40 % ont augmenté de taille.
Instabilité du génomeModifier
Les cancers sont connus pour présenter une instabilité du génome ou un phénotype mutateur. L’ADN codant pour les protéines à l’intérieur du noyau représente environ 1,5 % de l’ADN génomique total. Au sein de cet ADN codant pour les protéines (appelé exome), un cancer moyen du sein ou du côlon peut présenter environ 60 à 70 mutations altérant les protéines, dont environ 3 ou 4 peuvent être des mutations « pilotes » et les autres des mutations « passagères ». Cependant, le nombre moyen de mutations de la séquence d’ADN dans l’ensemble du génome (y compris les régions non codantes pour les protéines) dans un échantillon de tissu de cancer du sein est d’environ 20 000. Dans un échantillon moyen de tissu de mélanome (où les mélanomes ont une fréquence de mutation de l’exome plus élevée), le nombre total de mutations de séquence d’ADN est d’environ 80 000. Cela se compare à la très faible fréquence de mutation d’environ 70 nouvelles mutations dans l’ensemble du génome entre les générations (parent à enfant) chez les humains.
Les fréquences élevées de mutations dans les séquences nucléotidiques totales au sein des cancers suggèrent que souvent une altération précoce dans les défauts de champ donnant lieu à un cancer (par exemple, la zone jaune dans le diagramme de cette section) est une déficience dans la réparation de l’ADN. Facista et al. ont montré que les grands défauts de champ entourant les cancers du côlon (qui s’étendent jusqu’à environ 10 cm de chaque côté du cancer) présentent souvent des défauts épigénétiques dans 2 ou 3 protéines de réparation de l’ADN (ERCC1, XPF ou PMS2) dans toute la zone du défaut de champ. Les déficiences dans la réparation de l’ADN entraînent une augmentation des taux de mutation. Une déficience dans la réparation de l’ADN, en soi, peut permettre l’accumulation de dommages à l’ADN, et la synthèse de translesions sujets à des erreurs après certains de ces dommages peut donner lieu à des mutations. En outre, la réparation défectueuse de ces dommages accumulés à l’ADN peut donner lieu à des épimutations. Ces nouvelles mutations ou épimutations peuvent fournir un avantage prolifératif, générant un défaut de champ. Bien que les mutations/épimutations dans les gènes de réparation de l’ADN ne confèrent pas, en elles-mêmes, un avantage sélectif, elles peuvent être transportées en tant que passagers dans les cellules lorsque celles-ci acquièrent d’autres mutations/épimutations qui fournissent effectivement un avantage prolifératif.