Homo (von lateinisch homō, was ‚Mensch‘ bedeutet) ist die Gattung, die aus der ansonsten ausgestorbenen Gattung Australopithecus hervorging und die heutige Art Homo sapiens (moderner Mensch) sowie mehrere ausgestorbene Arten umfasst, die entweder als Vorfahren oder als eng verwandt mit dem modernen Menschen eingestuft werden (je nach Art), vor allem Homo erectus und Homo neanderthalensis.Die Gattung entstand mit dem Auftauchen des Homo habilis vor etwas mehr als zwei Millionen Jahren. Die Gattung Homo ist zusammen mit der Gattung Paranthropus wahrscheinlich Schwester von A. africanus in der Gattung Australopithecus, die sich ihrerseits zuvor von der Linie der Pan, den Schimpansen, abgespalten hatte.
Homo erectus erschien vor etwa zwei Millionen Jahren und verbreitete sich in mehreren frühen Wanderungen über Afrika (wo er als Homo ergaster bezeichnet wird) und Eurasien. Er war wahrscheinlich die erste menschliche Spezies, die in einer Jäger- und Sammlergesellschaft lebte und das Feuer beherrschte. Der Homo erectus war eine anpassungsfähige und erfolgreiche Spezies, die mehr als eine Million Jahre überlebte und sich vor etwa 500.000 Jahren allmählich in neue Arten aufspaltete.
Der Homo sapiens (anatomisch moderner Mensch) entstand vor etwa 300.000 bis 200.000 Jahren, wahrscheinlich in Afrika, und der Homo neanderthalensis entstand etwa zur gleichen Zeit in Europa und Westasien.H. sapiens breitete sich in mehreren Wellen aus Afrika aus, möglicherweise schon vor 250.000 Jahren, mit Sicherheit aber vor 130.000 Jahren, wobei die so genannte südliche Ausbreitung vor etwa 70-50.000 Jahren begann und zur dauerhaften Besiedlung Eurasiens und Ozeaniens vor 50.000 Jahren führte.Sowohl in Afrika als auch in Eurasien traf H. sapiens auf archaische Menschen und kreuzte sich mit ihnen. Es wird angenommen, dass separate archaische (nicht-sapiens) Menschenarten bis vor etwa 40.000 Jahren überlebt haben (Aussterben des Neandertalers), mit einem möglichen späten Überleben von Hybridarten bis vor 12.000 Jahren (Red Deer Cave People).
Namen und Taxonomie

Evolutionsbaumdiagramm mit Betonung der Unterfamilie Homininae und des Stammes Hominini. Nach der Abspaltung von der Linie zu den Ponginae spalteten sich die frühen Homininae in die Stämme Hominini und Gorillini. Die frühen Hominini spalteten sich weiter auf und trennten die Linie zu Homo von der Linie zu Pan. Gegenwärtig bezeichnet der Stamm Hominini die Unterstämme Hominina mit der Gattung Homo, Panina mit der Gattung Pan und Australopithecina mit mehreren ausgestorbenen Gattungen – die Unterstämme sind in dieser Grafik nicht beschriftet.
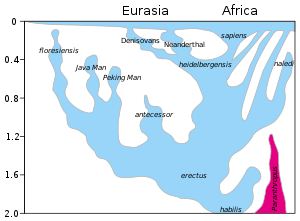
Ein Modell der Evolution der Gattung Homo über die letzten 2 Millionen Jahre (vertikale Achse). Die rasche „Out of Africa“-Expansion von H. sapiens ist oben im Diagramm angegeben, wobei die Vermischung mit Neandertalern, Denisovanern und nicht näher spezifizierten archaischen afrikanischen Homininen angezeigt wird. Das späte Überleben von robusten Australopithecinen (Paranthropus) neben Homo bis 1,2 Mya ist in lila dargestellt.
Hauptartikel: Taxonomie des Menschen, Namen für die menschlichen Arten Siehe Homininae für einen Überblick über die Taxonomie.
Das lateinische Substantiv homō (Genitiv hominis) bedeutet „Mensch“ oder „Mann“ im generischen Sinne von „Mensch, Menschheit“. Der binomiale Name Homo sapiens wurde von Carl Linnaeus (1758) geprägt. Namen für andere Arten der Gattung wurden ab der zweiten Hälfte des 19. Jahrhunderts eingeführt (H. neanderthalensis 1864, H. erectus 1892).
Auch heute ist die Gattung Homo nicht streng definiert. Seitdem die ersten menschlichen Fossilien langsam aus der Erde auftauchten, waren die Grenzen und Definitionen der Gattung Homo schlecht festgelegt und ständig im Fluss. Da es keinen Grund zu der Annahme gab, dass die Gattung jemals weitere Mitglieder haben würde, machte sich Carl Linnaeus nicht einmal die Mühe, Homo zu definieren, als er die Gattung im 18. Die Entdeckung des Neandertalers brachte die erste Ergänzung.
Die Gattung Homo erhielt ihren taxonomischen Namen, um anzudeuten, dass ihre Mitgliedsarten als menschlich eingestuft werden können. Und im Laufe der Jahrzehnte des 20. Jahrhunderts lieferten fossile Funde von vor- und frühmenschlichen Arten aus dem späten Miozän und frühen Pliozän eine reiche Mischung für die Diskussion von Klassifizierungen. Es gibt eine anhaltende Debatte über die Abgrenzung von Homo von Australopithecus – oder, in der Tat, über die Abgrenzung von Homo von Pan, da eine Gruppe von Wissenschaftlern argumentiert, dass die beiden Schimpansenarten eher in die Gattung Homo als in die Gattung Pan eingeordnet werden sollten. Trotzdem fällt die Klassifizierung der Fossilien von Homo mit den Beweisen für: (1) kompetenter menschlicher Bipedalismus bei Homo habilis, der vom früheren Australopithecus vor mehr als vier Millionen Jahren geerbt wurde, wie die Laetoli-Fußabdrücke zeigen; und (2) die menschliche Werkzeugkultur, die vor 2,5 Millionen Jahren begann.
Vom späten 19. bis zur Mitte des 20. Jahrhunderts wurden eine Reihe neuer taxonomischer Namen, einschließlich neuer Gattungsnamen, für frühe menschliche Fossilien vorgeschlagen; die meisten wurden seitdem mit Homo zusammengelegt, in Anerkennung der Tatsache, dass Homo erectus eine einzige und singuläre Art mit einer großen geographischen Verbreitung früher Wanderungen war. Viele dieser Namen werden nun als „Synonyme“ mit Homo bezeichnet, darunter Pithecanthropus, Protanthropus, Sinanthropus, Cyphanthropus, Africanthropus, Telanthropus, Atlanthropus und Tchadanthropus.
Die Klassifizierung der Gattung Homo in Arten und Unterarten unterliegt unvollständigen Informationen und ist nach wie vor schlecht gemacht. Dies hat dazu geführt, dass selbst in wissenschaftlichen Arbeiten gebräuchliche Namen („Neandertaler“ und „Denisovaner“) verwendet werden, um trinomiale Namen oder die Mehrdeutigkeit der Klassifizierung von Gruppen als incertae sedis (unsichere Platzierung) zu vermeiden – zum Beispiel H. neanderthalensis vs. H. sapiens neanderthalensis oder H. georgicus vs. H. erectus georgicus. Einige kürzlich ausgestorbene Arten der Gattung Homo sind erst kürzlich entdeckt worden und haben noch keine einheitlichen binomischen Namen (siehe Denisova-Hominin und Red Deer Cave People). Seit Beginn des Holozäns ist es wahrscheinlich, dass Homo sapiens (anatomisch moderner Mensch) die einzige existierende Art von Homo ist.
John Edward Gray (1825) war ein früher Verfechter der Klassifizierung von Taxa durch die Bezeichnung von Stämmen und Familien.Wood und Richmond (2000) schlugen vor, die Hominini („Homininen“) als einen Stamm zu bezeichnen, der alle Arten der frühen Menschen und Vormenschen umfasst, die bis nach dem letzten gemeinsamen Vorfahren von Schimpanse und Mensch zurückreichen; und dass Hominina als Unterstamm von Hominini bezeichnet wird, um nur die Gattung Homo zu umfassen – das heißt, ohne die früheren aufrecht gehenden Homininen des Pliozäns wie Australopithecus, Orrorin tugenensis, Ardipithecus oder Sahelanthropus.Alternative Bezeichnungen zu Hominina existierten bzw. wurden angeboten: Australopithecinae (Gregory & Hellman 1939) und Preanthropinae (Cela-Conde & Altaba 2002);und später schlugen Cela-Conde und Ayala (2003) vor, die vier Gattungen Australopithecus, Ardipithecus, Praeanthropus und Sahelanthropus mit Homo innerhalb der Hominini (sans pan) zu gruppieren.
Evolution
Weitere Informationen: Zeitleiste der menschlichen Evolution Siehe Hominini und Schimpanse-Mensch letzter gemeinsamer Vorfahre für die Trennung von Australopithecina und Panina.
Australopithecus
Weitere Informationen: Australopithecus
Forensische Rekonstruktion von A. afarensis
Einige Arten, darunter Australopithecus garhi, Australopithecus sediba, Australopithecus africanus und Australopithecus afarensis, sind als direkter Vorfahre oder Schwester der Homo-Linie vorgeschlagen worden.Diese Arten haben morphologische Merkmale, die sie mit Homo in Einklang bringen, aber es gibt keinen Konsens darüber, aus welcher Art Homo hervorging.
Vor allem seit den 2010er Jahren ist die Abgrenzung von Homo zu Australopithecus umstrittener geworden. Traditionell wird das Aufkommen von Homo mit dem ersten Gebrauch von Steinwerkzeugen (der Oldowan-Industrie) und damit per Definition mit dem Beginn des Jungpaläolithikums gleichgesetzt.
Im Jahr 2010 wurden jedoch Beweise vorgelegt, die Australopithecus afarensis den Gebrauch von Steinwerkzeugen vor etwa 3,3 Millionen Jahren zuzuschreiben scheinen, also fast eine Million Jahre vor dem ersten Auftreten von Homo.
LD 350-1, ein fossiles Unterkieferfragment, das auf 2.8 Mya, das 2015 in Afar, Äthiopien, entdeckt wurde, wurde als eine Kombination von „primitiven Merkmalen, die bei frühem Australopithecus gesehen wurden, mit abgeleiteter Morphologie, die bei späterem Homo beobachtet wurde“ beschrieben.
Einige Autoren würden die Entwicklung von Homo nahe an oder sogar über 3 Mya hinausschieben.
Andere haben Zweifel geäußert, ob Homo habilis in Homo eingeschlossen werden sollte, und schlagen stattdessen einen Ursprung von Homo mit Homo erectus bei etwa 1,9 Mya vor.
Die auffälligste physiologische Entwicklung zwischen den früheren australopithecinen Arten und Homo ist die Zunahme des endokranialen Volumens (ECV), von etwa 460 cm3 (28 cu in) bei A. garhi auf 660 cm3 (40 cu in) bei H. habilis und weiter auf 760 cm3 (46 cu in) bei H. erectus, 1.250 cm3 (76 cu in) bei H. heidelbergensis und bis zu 1.760 cm3 (107 cu in) bei H. neanderthalensis.Ein stetiger Anstieg des Schädelvolumens ist jedoch bereits bei Autralopithecina zu beobachten und endet nicht nach der Entstehung von Homo, so dass es nicht als objektives Kriterium zur Definition der Entstehung der Gattung dienen kann.
Homo habilis
Forensische Rekonstruktion von Homo habilis, Exponat im LWL-Museum für Archäologie, Herne, Deutschland (Foto 2007).
Homo habilis entstand etwa 2,1 Mya. Schon vor 2010 gab es Vorschläge, dass H. habilis nicht außerhalb der Gattung Homo in den weiteren Australopithecus gestellt werden sollte.Der Hauptgrund, H. habilis in Homo einzuschließen, sein unbestrittener Werkzeuggebrauch, ist mit der Entdeckung des Werkzeuggebrauchs von Australopithecus mindestens eine Million Jahre vor H. habilis obsolet geworden. Außerdem wurde H. habilis lange Zeit als Vorfahre des grazilen Homo ergaster (Homo erectus) angesehen. 2007 wurde entdeckt, dass H. habilis und H. erectus für eine beträchtliche Zeit koexistierten, was darauf hindeutet, dass H. erectus nicht unmittelbar von H. habilis, sondern von einem gemeinsamen Vorfahren abstammt.2013 wurde mit der Veröffentlichung des Dmanisi-Schädels 5 weniger sicher, dass der asiatische H. erectus ein Nachkomme des afrikanischen H. ergaster ist, der wiederum von H. habilis abstammt.Stattdessen scheinen H. ergaster und H. erectus Varianten derselben Spezies zu sein, die entweder in Afrika oder Asien entstanden sein könnten und sich um 0,5 Mya in ganz Eurasien (einschließlich Europa, Indonesien, China) verbreitet haben.
Homo erectus
Hauptartikel: Homo erectus
Homo erectus ist oft angenommen worden, dass er sich anagenetisch aus Homo habilis vor etwa 2 Millionen Jahren entwickelt hat.
Dieses Szenario wurde durch die Entdeckung von Homo erectus georgicus, frühen Exemplaren von H. erectus, die im Kaukasus gefunden wurden und die Übergangsmerkmale mit H. habilis aufzuweisen schienen, gestärkt. Da die frühesten Belege für H. erectus außerhalb Afrikas gefunden wurden, wurde es als plausibel erachtet, dass sich H. erectus in Eurasien entwickelte und dann nach Afrika zurückwanderte. Basierend auf Fossilien aus der Koobi Fora Formation, östlich des Turkana Sees in Kenia, argumentierten Spoor et al. (2007), dass H. habilis über die Entstehung von H. erectus hinaus überlebt haben könnte, so dass die Evolution von H. erectus nicht anagenetisch gewesen wäre und H. erectus neben H. habilis für etwa eine halbe Million Jahre (vor 1,9 bis 1,4 Millionen Jahren), während des frühen Kalabriums.
Eine separate südafrikanische Art Homo gautengensis wurde 2010 als zeitgleich mit Homo erectus postuliert.
Phylogenie
Eine Taxonomie des Homo innerhalb der Menschenaffen wird wie folgt eingeschätzt, wobei Paranthropus und Homo innerhalb von Australopithecus auftauchen (hier kladistisch dargestellt mit Paranthropus, Kenyanthropus und Homo). Die genaue Phylogenie innerhalb von Australopithecus ist noch sehr umstritten. Ungefähre Radiationsdaten der Tochterkladen sind in Millionen von Jahren vor (Mya) angegeben. Graecopithecus, Sahelanthropus, Orrorin, möglicherweise Schwestern von Australopithecus, sind hier nicht dargestellt. Beachten Sie, dass die Benennung der Gruppierungen manchmal durcheinander gerät, da oft bestimmte Gruppierungen vermutet werden, bevor eine kladistische Analyse durchgeführt wird.
| Hominoidea |
|
||||||||||||||||||||||||
| (20.4 Mya) |
| Australopithecinen |
Einige der Homo-Linien scheinen durch Introgression in andere Linien Nachkommenschaft überlebt zu haben. Eine archaische Linie, die sich vor 1,5 Millionen Jahren von den anderen menschlichen Linien trennte, vielleicht H. erectus, könnte sich vor etwa 55.000 Jahren in die Denisovaner eingekreuzt haben. Homo erectus s.s. überlebte bis vor 27.000 Jahren, und der noch basalere Homo florensiensis überlebte bis vor 50.000 Jahren. Außerdem ähnelt ein auf 14.000 Jahre datierter Oberschenkelknochen, der in einer Maludong-Höhle (Red Deer Cave people) gefunden wurde, sehr alten Arten wie dem frühen Homo erectus oder der noch archaischeren Linie, dem Homo habilis, der vor etwa 1,5 Millionen Jahren lebte. Ein Teil der 1,5 Millionen Jahre alten Homo erectus-ähnlichen Linie scheint über die Denisovaner und speziell über die Papuas und die australischen Ureinwohner in den modernen Menschen eingedrungen zu sein. Es gibt Hinweise auf eine Introgression von H. Heidelbergensis in H. sapiens. Die Genome von Menschen, die nicht aus Afrika südlich der Sahara stammen, zeigen zahlreiche unabhängige Introgressionsereignisse, an denen Neandertaler und in einigen Fällen auch Denisovaner vor etwa 45.000 Jahren beteiligt waren. Ebenso scheint die genetische Struktur der Subsahara-Afrikaner auf eine Introgression aus einer bestimmten, noch nicht identifizierten archaischen Menschenlinie wie H. heidelbergensis hinzudeuten. Australopithecus sediba soll aufgrund seiner Position in Bezug auf z.B. Homo habilis und Homo floresiensis in Homo sediba umbenannt werden. DispersalSiehe auch: Frühe menschliche Expansionen aus Afrika heraus, Vermischung des archaischen Menschen mit dem modernen Menschen, Frühe menschliche Migrationen Vor etwa 1,8 Millionen Jahren ist Homo erectus sowohl in Ostafrika (Homo ergaster) als auch in Westasien (Homo georgicus) vertreten. Die Vorfahren des indonesischen Homo floresiensis haben Afrika möglicherweise noch früher verlassen.  Sukzessive Ausbreitung von Homo erectus (gelb), Homo neanderthalensis (ocker) und Homo sapiens (rot). Homo erectus und verwandte oder abgeleitete archaische Menschenarten verbreiten sich in den nächsten 1,5 Millionen Jahren über Afrika und Eurasien (siehe: Rezentes Afrika als Ursprung des modernen Menschen). Europa wird um etwa 0,5 Mya von Homo heidelbergensis erreicht. Homo neanderthalensis und Homo sapiens entwickeln sich nach etwa 300 kya. Homo naledi ist um 300 kya im südlichen Afrika präsent. H. sapiens breitete sich bald nach seinem ersten Auftauchen in mehreren Wellen über ganz Afrika und nach Westasien aus, möglicherweise schon ab 250 kya, sicher aber ab 130 kya. Im Juli 2019 meldeten Anthropologen die Entdeckung von 210.000 Jahre alten Überresten eines H. sapiens und 170.000 Jahre alten Überresten eines H. neanderthalensis in der Apidima-Höhle auf dem Peloponnes in Griechenland, die mehr als 150.000 Jahre älter sind als bisherige H. sapiens-Funde in Europa. Besonders bemerkenswert ist die südliche Ausbreitung von H. sapiens um 60 kya, die zur dauerhaften Besiedlung Ozeaniens und Eurasiens durch anatomisch moderne Menschen führte. H. sapiens kreuzte sich sowohl in Afrika als auch in Eurasien mit archaischen Menschen, in Eurasien vor allem mit Neandertalern und Denisovanen. Unter den heute lebenden Populationen des Homo sapiens findet sich die tiefste zeitliche Trennung bei den San im südlichen Afrika, die auf etwa 130.000 Jahre oder möglicherweise mehr als 300.000 Jahre geschätzt wird. Die zeitliche Trennung bei den Nicht-Afrikanern liegt in der Größenordnung von 60.000 Jahren bei den Australo-Melanesiern.Die Aufteilung der Europäer und Ostasiaten liegt in der Größenordnung von 50.000 Jahren, mit wiederholten und bedeutenden Vermischungsereignissen in ganz Eurasien während des Holozäns. Archäische Menschenarten haben möglicherweise bis zum Beginn des Holozäns überlebt (Red Deer Cave People), obwohl sie größtenteils ausgestorben sind oder von den expandierenden H. sapiens-Populationen um 40 kya absorbiert wurden (Aussterben des Neandertalers). Liste der LinienSiehe auch: Liste der Fossilien der menschlichen Evolution Der Artstatus von H. rudolfensis, H. ergaster, H. georgicus, H. antecessor, H. cepranensis, H. rhodesiensis, H. neanderthalensis, Denisova-Hominin, Red Deer Cave People und H. floresiensis bleibt umstritten. H. heidelbergensis und H. neanderthalensis sind eng miteinander verwandt und wurden als Unterarten von H. sapiens angesehen. Es gab in der Vergangenheit einen Trend, „neue menschliche Spezies“ zu postulieren, die nur auf einem einzelnen Fossil basierten. Ein „minimalistischer“ Ansatz zur Taxonomie des Menschen erkennt höchstens drei Arten an, Homo habilis (2,1-1,5 Mya, Zugehörigkeit zu Homo fraglich), Homo erectus (1,8-0,1 Mya, einschließlich der Mehrheit des Alters der Gattung und der Mehrheit der archaischen Varianten als Unterarten, einschließlich H. heidelbergensis als späte oder Übergangsvariante) und Homo sapiens (300 kya bis heute, einschließlich H. neanderthalensis und anderer Varianten als Unterarten). „Spezies“ bedeutet in diesem Zusammenhang nicht unbedingt, dass Hybridisierung und Introgression zu diesem Zeitpunkt unmöglich waren. Er wird jedoch oft als bequemer Begriff verwendet, sollte aber im besten Fall als eine generische Linie und im schlimmsten Fall als Cluster verstanden werden. Im Allgemeinen sind die Definitionen und die Methodik der Abgrenzungskriterien für „Arten“ in der Anthropologie oder Paläontologie nicht allgemein anerkannt. In der Tat können sich Säugetiere und typischerweise 2 bis 3 Millionen Jahre oder länger kreuzen, so dass alle zeitgenössischen Homo-„Arten“ der Gattung Homo potenziell in der Lage gewesen wären, sich zu dieser Zeit zu kreuzen, und eine Introgression von außerhalb der Gattung Homo kann nicht a priori ausgeschlossen werden. Es wurde vorgeschlagen, dass H. naledi eine Kreuzung mit einem spät überlebenden Australipithen (gemeint ist: jenseits von Homo, Anm. d. Red.) gewesen sein könnte, trotz der Tatsache, dass diese Linien allgemein als längst ausgestorben gelten. Wie oben besprochen, haben viele Introgressionen zwischen den Linien stattgefunden, mit Beweisen für Introgression nach einer Trennung von 1,5 Millionen Jahren. Fußnoten
|
