Homo (z łac. homō, co oznacza „człowiek”) to rodzaj, który powstał z wymarłego w innym przypadku rodzaju Australopithecus, który obejmuje wymarły gatunek Homo sapiens (współczesny człowiek), plus kilka wymarłych gatunków sklasyfikowanych jako przodkowie lub blisko spokrewnieni ze współczesnymi ludźmi (w zależności od gatunku), zwłaszcza Homo erectus i Homo neanderthalensis.Przyjmuje się, że rodzaj powstał wraz z pojawieniem się Homo habilis, nieco ponad dwa miliony lat temu. Rodzaj Homo, wraz z rodzajem Paranthropus jest prawdopodobnie siostrą A. africanus w rodzaju Australopithecus, który sam wcześniej oddzielił się od linii Pan, szympansów.
Homo erectus pojawił się około dwóch milionów lat temu i w kilku wczesnych migracjach rozprzestrzenił się w całej Afryce (gdzie został nazwany Homo ergaster) i Eurazji. Był to prawdopodobnie pierwszy gatunek ludzki, który żył w społeczeństwie łowców-zbieraczy i kontrolował ogień. Jako gatunek przystosowawczy i odnoszący sukcesy, Homo erectus przetrwał ponad milion lat i stopniowo rozdzielił się na nowe gatunki około 500 000 lat temu.
Homo sapiens (anatomicznie współcześni ludzie) pojawił się blisko 300 000 do 200 000 lat temu, najprawdopodobniej w Afryce, a Homo neanderthalensis pojawił się mniej więcej w tym samym czasie w Europie i Azji Zachodniej.H. sapiens rozproszył się z Afryki w kilku falach, od prawdopodobnie już 250.000 lat temu, a na pewno do 130.000 lat temu, tak zwana południowa dyspersja rozpoczynająca się około 70-50.000 lat temu, prowadząca do trwałej kolonizacji Eurazji i Oceanii do 50.000 lat temu.Zarówno w Afryce, jak i Eurazji, H. sapiens spotkał się i krzyżował z archaicznymi ludźmi. Uważa się, że oddzielne archaiczne (nie-sapiens) gatunki ludzkie przetrwały do około 40 000 lat temu (wymarcie neandertalczyków), z możliwym późnym przetrwaniem gatunków hybrydowych aż do 12 000 lat temu (ludzie z jaskini Red Deer).
Nazwy i taksonomia

Wykres drzewa ewolucyjnego podkreślający podrodzinę Homininae i plemię Hominini. Po odłączeniu się od linii do Ponginae, wczesne Homininae podzieliły się na plemiona Hominini i Gorillini. Wczesne Hominini uległy dalszemu podziałowi, oddzielając linię do Homo od linii do Pan. Obecnie plemię Hominini wyznacza podplemię Hominina, zawierające rodzaj Homo; Panina, rodzaj Pan; oraz Australopithecina, z kilkoma wymarłymi rodzajami – podplemiona nie są oznaczone na tym wykresie.
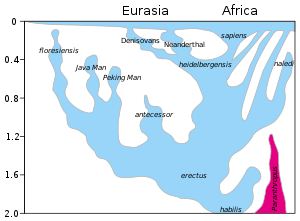
Model ewolucji rodzaju Homo w ciągu ostatnich 2 milionów lat (oś pionowa). Szybki „Out of Africa” ekspansja H. sapiens jest wskazany w górnej części wykresu, z domieszką wskazano z Neandertalczyków, Denisovans i nieokreślonych archaicznych homininów afrykańskich. Późne przetrwanie wytrzymałych australopiteki (Paranthropus) obok Homo do 1,2 Mya jest zaznaczone na fioletowo.
Główne artykuły: Taksonomia człowieka, Nazwy dla gatunku ludzkiego Zobacz Homininae dla przeglądu taksonomii.
Łaciński rzeczownik homō (genitive hominis) oznacza „istotę ludzką” lub „człowieka” w znaczeniu ogólnym „istota ludzka, rodzaj ludzki”. Nazwa binominalna Homo sapiens została ukuta przez Carla Linneusza (1758). Nazwy dla innych gatunków z tego rodzaju wprowadzono począwszy od drugiej połowy XIX wieku (H. neanderthalensis 1864, H. erectus 1892).
Do dziś rodzaj Homo nie jest ściśle zdefiniowany. Odkąd wczesny ludzki zapis kopalny zaczął powoli wyłaniać się z ziemi, granice i definicje rodzaju Homo były słabo zdefiniowane i stale się zmieniały. Ponieważ nie było powodu, by sądzić, że rodzaj ten kiedykolwiek będzie miał dodatkowych członków, Carl Linneusz nie zadał sobie nawet trudu, by zdefiniować Homo, gdy po raz pierwszy stworzył go dla ludzi w XVIII wieku. Odkrycie neandertalczyka przyniosło pierwszy dodatek.
Rodzaj Homo otrzymał swoją nazwę taksonomiczną, aby zasugerować, że jego gatunki członkowskie mogą być klasyfikowane jako ludzkie. A w ciągu kilkudziesięciu lat XX wieku znaleziska kopalne przedludzkich i wczesnych gatunków ludzkich z późnego miocenu i wczesnego pliocenu dostarczyły bogatej mieszanki do dyskusji nad klasyfikacjami. Wciąż toczy się debata na temat rozgraniczenia Homo od Australopithecus – lub, w rzeczy samej, rozgraniczenia Homo od Pan, jako że jedna grupa naukowców twierdzi, że te dwa gatunki szympansów powinny być klasyfikowane do rodzaju Homo, a nie Pan. Mimo to, klasyfikowanie skamieniałości Homo zbiega się z dowodami na: (1) kompetentnego ludzkiego dwunożności u Homo habilis odziedziczonego po wcześniejszym Australopithecus sprzed ponad czterech milionów lat, jak pokazują odciski stóp Laetoli; i (2) ludzkiej kultury narzędziowej rozpoczętej przez 2,5 miliona lat temu.
Od końca XIX do połowy XX wieku zaproponowano szereg nowych nazw taksonomicznych, w tym nowych nazw rodzajowych, dla skamieniałości wczesnego człowieka; większość z nich została od tego czasu połączona z Homo w uznaniu, że Homo erectus był pojedynczym gatunkiem o dużym zasięgu geograficznym wczesnych migracji. Wiele takich nazw jest teraz dubbingowane jako „synonimy” z Homo, w tym Pithecanthropus, Protanthropus, Sinanthropus, Cyphanthropus, Africanthropus, Telanthropus, Atlanthropus i Tchadanthropus.
Klasyfikacja rodzaju Homo na gatunki i podgatunki jest przedmiotem niepełnych informacji i pozostaje słabo zrobione. Doprowadziło to do używania wspólnych nazw („Neandertalczyk” i „Denisowianin”), nawet w pracach naukowych, aby uniknąć nazw trójczłonowych lub niejednoznaczności klasyfikacji grup jako incertae sedis (niepewne umiejscowienie) – na przykład H. neanderthalensis vs. H. sapiens neanderthalensis, lub H. georgicus vs. H. erectus georgicus. Niektóre niedawno wymarłe gatunki z rodzaju Homo zostały dopiero niedawno odkryte i nie mają jeszcze zgodnych nazw binomialnych (zob. Denisova hominin i Red Deer Cave people). Od początku holocenu, prawdopodobnie Homo sapiens (anatomicznie współczesny człowiek) jest jedynym wymarłym gatunkiem Homo.
John Edward Gray (1825) był wczesnym zwolennikiem klasyfikacji taksonów poprzez wyznaczanie plemion i rodzin.Wood i Richmond (2000) zaproponowali, by Hominini („homininy”) określić jako plemię, które obejmowałoby wszystkie gatunki wczesnych ludzi i prehumanów przodków ludzi, aż do okresu po ostatnim wspólnym przodku szympansa i człowieka; oraz aby Hominina została określona jako podplemię Hominini i obejmowała jedynie rodzaj Homo – to znaczy nie obejmowała wcześniejszych, chodzących na stojąco homininów z Pliocenu, takich jak Australopithecus, Orrorin tugenensis, Ardipithecus czy Sahelanthropus.Istniały lub były proponowane nazwy alternatywne do Hominina: Australopithecinae (Gregory & Hellman 1939) i Preanthropinae (Cela-Conde & Altaba 2002);a później Cela-Conde i Ayala (2003) zaproponowali, by cztery rodzaje Australopithecus, Ardipithecus, Praeanthropus i Sahelanthropus zgrupować z Homo w ramach Hominini (sans pan).
Ewolucja
Dalsze informacje: Oś czasu ewolucji człowieka Zobacz Hominini i Szympans-człowiek ostatni wspólny przodek dla oddzielenia Australopithecina i Panina.
Australopithecus
Dalsze informacje: Australopithecus
Forensic reconstruction of A. afarensis
Kilka gatunków, w tym Australopithecus garhi, Australopithecus sediba, Australopithecus africanus i Australopithecus afarensis, zostało zaproponowanych jako bezpośredni przodek lub siostra linii Homo.Gatunki te mają cechy morfologiczne, które łączą je z Homo, ale nie ma zgody co do tego, który z nich dał początek Homo.
Szczególnie od lat 2010-tych, rozgraniczenie Homo od Australopithecus stało się bardziej kontrowersyjne. Tradycyjnie przyjmuje się, że pojawienie się Homo zbiegło się z pierwszym użyciem narzędzi kamiennych (przemysł Oldowan), a więc z definicji z początkiem dolnego paleolitu.Ale w 2010 r. przedstawiono dowody, które wydają się przypisywać Australopithecus afarensis użycie narzędzi kamiennych około 3,3 mln lat temu, blisko milion lat przed pierwszym pojawieniem się Homo.LD 350-1, fragment kopalnej żuchwy datowany na 2.8 Mya, odkryty w 2015 r. w Afar w Etiopii, został opisany jako łączący „prymitywne cechy widoczne u wczesnego Australopithecus z pochodną morfologią obserwowaną u późniejszego Homo.Niektórzy autorzy przesunęliby rozwój Homo blisko lub nawet poza 3 Mya.Inni wyrazili wątpliwości, czy Homo habilis powinien być włączony do Homo, proponując zamiast tego pochodzenie Homo z Homo erectus w przybliżeniu 1,9 Mya.
Najważniejszym rozwojem fizjologicznym między wcześniejszymi australopitecznymi gatunkami a Homo jest wzrost objętości wewnątrzczaszkowej (ECV), z około 460 cm3 (28 cu in) u A. garhi do 660 cm3 (40 cu in) u H. habilis i dalej do 760 cm3 (46 cu in) u H. erectus, 1.250 cm3 (76 cu in) u H. heidelbergensis i do 1.760 cm3 (107 cu in) u H. neanderthalensis.Stały wzrost pojemności czaszki obserwowany jest jednak już u Autralopithecina i nie kończy się po powstaniu Homo, nie służy więc jako obiektywne kryterium definiujące powstanie rodzaju.
Homo habilis
Forensyczna rekonstrukcja Homo habilis, eksponat w LWL-Museum für Archäologie, Herne, Niemcy (zdjęcie z 2007 r.).
Homo habilis pojawił się około 2,1 Mya. Już przed 2010 r. pojawiły się sugestie, że H. habilis nie powinien być umieszczany poza rodzajem Homo w szerszym Australopithecus.Główny powód włączenia H. habilis do Homo, jego bezsporne użycie narzędzi, stał się nieaktualny wraz z odkryciem użycia narzędzi Australopithecus co najmniej milion lat przed H. Co więcej, H. habilis był długo uważany za przodka bardziej zgrabnego Homo ergaster (Homo erectus).W 2007 r. odkryto, że H. habilis i H. erectus współistniały przez dłuższy czas, co sugeruje, że H. erectus nie pochodzi bezpośrednio od H. habilis, ale zamiast tego od wspólnego przodka.Wraz z publikacją czaszki Dmanisi 5 w 2013 roku, stało się mniej pewne, że azjatycki H. erectus jest potomkiem afrykańskiego H. ergaster, który z kolei pochodził od H. habilis.Zamiast tego, H. ergaster i H. erectus wydają się być wariantami tego samego gatunku, który mógł pochodzić z Afryki lub Azji i szeroko rozproszyć się po Eurazji (w tym Europie, Indonezji, Chinach) do 0,5 Mya.
Homo erectus
Główne artykuły: Homo erectus
Homo erectus często był zakładany, że rozwinął się anagenetycznie z Homo habilis od około 2 milionów lat temu.Ten scenariusz został wzmocniony wraz z odkryciem Homo erectus georgicus, wczesnych okazów H. erectus znalezionych na Kaukazie, które wydawały się wykazywać cechy przejściowe z H. habilis. Ponieważ najwcześniejsze dowody na H. erectus znaleziono poza Afryką, uznano za prawdopodobne, że H. erectus rozwinął się w Eurazji, a następnie migrował z powrotem do Afryki. Na podstawie skamieniałości z formacji Koobi Fora, na wschód od jeziora Turkana w Kenii, Spoor et al. (2007) argumentował, że H. habilis mógł przetrwać poza pojawieniem się H. erectus, tak że ewolucja H. erectus nie byłaby anagenetyczna, a H. erectus istniałby obok H. habilis przez około pół miliona lat (1,9 do 1,4 mln lat temu), podczas wczesnego kalabryjskiego.
Odrębny południowoafrykański gatunek Homo gautengensis został postulowany jako współczesny Homo erectus w 2010 roku.
Phylogenezy
Taksonomia Homo w obrębie małp człekokształtnych jest oceniana następująco, z Paranthropus i Homo wyłaniającymi się w obrębie Australopithecus (pokazany tutaj kladystycznie przyznając Paranthropus, Kenyanthropus i Homo). Dokładna filogeneza w obrębie Australopithecus jest nadal wysoce kontrowersyjna. Przybliżone daty radiacji kladów pochodnych podano w milionach lat temu (Mya). Graecopithecus, Sahelanthropus, Orrorin, prawdopodobnie siostry Australopithecus, nie są tu pokazane. Zauważmy, że nazewnictwo grup jest czasem niejasne, gdyż często pewne grupy są zakładane przed wykonaniem analizy kladystycznej.
| Hominoidea |
|
||||||||||||||||||||||||
| (20.4 Mya) |
| Australopiteki |
Kilka z linii Homo wydaje się mieć potomstwo, które przetrwało dzięki introgresji do innych linii. Archaiczna linia oddzielająca się od innych ludzkich linii 1,5 miliona lat temu, być może H. erectus, mogła wmieszać się w Denisovans około 55 000 lat temu. Homo erectus s.s. przetrwał do 27.000 lat temu, a jeszcze bardziej bazalny Homo florensiensis przetrwał do 50.000 lat temu. Co więcej, kość udowa, datowana na 14 000 lat, znaleziona w jaskini Maludong (ludzie z Red Deer Cave) silnie przypomina bardzo starożytne gatunki, takie jak wczesny Homo erectus lub jeszcze bardziej archaiczny ród Homo habilis, który żył około 1,5 miliona lat temu. Niektóre z 1,5 miliona lat Homo erectus-jak linii wydaje się, że uczynił swoją drogę do współczesnych ludzi przez Denisovans, a zwłaszcza do Papuasów i rdzennych Australijczyków. Istnieją dowody na introgresję H. Heidelbergensis do H. sapiens. Genomy ludzi spoza Afryki Subsaharyjskiej pokazują to, co wydaje się być liczne niezależne wydarzenia introgresji z udziałem Neandertalczyków, a w niektórych przypadkach także Denisowian około 45.000 lat temu. Podobnie struktura genetyczna mieszkańców Afryki Subsaharyjskiej wydaje się wskazywać na introgresję z odrębnej, jak dotąd niezidentyfikowanej archaicznej linii ludzkiej, takiej jak H. heidelbergensis. Australopithecus sediba jest gotowy do zmiany nazwy na Homo sediba ze względu na jego pozycję w stosunku do np. Homo habilis i Homo floresiensis. RozproszenieZobacz także: Wczesne ekspansje człowieka poza Afrykę, Domieszki człowieka archaicznego z człowiekiem współczesnym, Wczesne migracje człowieka Do około 1,8 mln lat temu Homo erectus jest obecny zarówno w Afryce Wschodniej (Homo ergaster), jak i w Azji Zachodniej (Homo georgicus).Przodkowie indonezyjskiego Homo floresiensis mogli opuścić Afrykę jeszcze wcześniej.  Sukcesywne rozprzestrzenianie się Homo erectus (żółty), Homo neanderthalensis (ochra) i Homo sapiens (czerwony). Homo erectus i pokrewne lub pochodne archaiczne gatunki ludzkie przez następne 1,5 mln lat rozprzestrzeniały się po Afryce i Eurazji (patrz: Ostatnie afrykańskie pochodzenie współczesnych ludzi). Do Europy dociera Homo heidelbergensis około 0,5 Mya. Homo neanderthalensis i Homo sapiens rozwijają się po około 300 kya. Homo naledi jest obecny w południowej Afryce do 300 kya. H. sapiens wkrótce po pierwszym pojawieniu się rozprzestrzenił się w całej Afryce, a także w Azji Zachodniej w kilku falach, być może już w 250 kya, a na pewno do 130 kya. W lipcu 2019 r. antropolodzy poinformowali o odkryciu 210 000-letnich szczątków H. sapiens i 170 000-letnich szczątków H. neanderthalensis w jaskini Apidima, Peloponez, Grecja, ponad 150 000 lat starszych niż poprzednie znaleziska H. sapiens w Europie. Najbardziej godna uwagi jest Południowa Dyspersja H. sapiens około 60 kya, która doprowadziła do trwałego zaludnienia Oceanii i Eurazji przez anatomicznie nowoczesnych ludzi. H. sapiens krzyżował się z archaicznymi ludźmi zarówno w Afryce, jak i w Eurazji, w Eurazji zwłaszcza z neandertalczykami i denisowianami. Wśród istniejących populacji Homo sapiens, najgłębszy podział czasowy występuje u ludu San z południowej Afryki, szacowany na blisko 130 000 lat, lub prawdopodobnie ponad 300 000 lat temu.Podział czasowy wśród nie-Afrykanów jest rzędu 60 000 lat w przypadku Australo-Melanesians.Podział Europejczyków i wschodnich Azjatów jest rzędu 50 000 lat, z powtarzającymi się i znaczącymi domieszkami w całej Eurazji podczas holocenu. Archaiczne gatunki ludzkie mogły przetrwać do początku holocenu (ludzie z Red Deer Cave), choć w większości wymarły lub zostały wchłonięte przez rozszerzające się populacje H. sapiens do 40 kya (wymieranie neandertalczyków). Lista liniiZobacz także: Lista skamieniałości ewolucji człowieka Status gatunkowy H. rudolfensis, H. ergaster, H. georgicus, H. antecessor, H. cepranensis, H. rhodesiensis, H. neanderthalensis, Denisova hominin, Red Deer Cave people i H. floresiensis pozostaje dyskusyjny. H. heidelbergensis i H. neanderthalensis są blisko spokrewnione ze sobą i zostały uznane za podgatunki H. sapiens. W przeszłości istniała tendencja do postulowania „nowych gatunków człowieka” w oparciu o tak niewiele, jak pojedyncza skamielina. Minimalistyczne” podejście do taksonomii człowieka uznaje co najwyżej trzy gatunki, Homo habilis (2.1-1.5 Mya, przynależność do Homo wątpliwa), Homo erectus (1.8-0.1 Mya, obejmujący większość wieku rodzaju, oraz większość archaicznych odmian jako podgatunki, w tym H. heidelbergensis jako odmianę późną lub przejściową) i Homo sapiens (300 kya do dziś, w tym H. neanderthalensis i inne odmiany jako podgatunki). „Gatunek” nie musi w tym kontekście oznaczać, że hybrydyzacja i introgresja były wówczas niemożliwe. Jest jednak często używany jako wygodny termin, ale należy go traktować jako oznaczający w najlepszym przypadku linię rodzajową, a w najgorszym skupiska. W ogóle definicje i metodologia kryteriów wyodrębniania „gatunków” nie są powszechnie uzgodnione w antropologii czy paleontologii. Istotnie, ssaki i mogą się zazwyczaj krzyżować przez 2 do 3 milionów lat lub dłużej, więc wszystkie współczesne „gatunki” Homo w rodzaju Homo potencjalnie byłyby w stanie krzyżować się w tym czasie, a introgresja spoza rodzaju Homo nie może być a priori wykluczona. Sugeruje się, że H. naledi mógł być hybrydą z późno żyjącym australipitekem (rozumianym jako poza Homo, red.), mimo że te linie generalnie uważane są za dawno wymarłe. Jak omówiono powyżej, wiele introgresji wystąpiło między liniami, z dowodami na introgresję po oddzieleniu 1,5 miliona lat. Przypisy
|
